One of the best things about working at a university your whole life is the amazing talks and speakers that you get to see. I’ve just been to see Richard Andersen from Caltech talking on the ‘Parietal lobe in action’ (Parietal lobe and action = two of my favourite things..). Suffice to say, it was impressive. Here’s a little summary:
Posterior parietal cortex (PPC) is association cortex (i.e. not primary cortex) lying between primary somatosensory cortex and visual cortex.
In 1909 Balint reported a case of a patient with bilateral damage to PPC which resulted in deficits in eye movements, reaching movements (a condition known as optic ataxia) and spatial attention (known as spatial neglect). This combination of symptoms went on to be known as Balint's syndrome. He concluded that PPC was central to sensory motor transformations and that damage to this area resulted in deficits in processes that required such transformations from sensory input to motor output.
Then in 1918, Holmes reported a case of patients with similar bilateral PPC lesions but he attributed their deficits to a purely visual deficit, rather than a sensory-motor deficit.
In 1988 Perenin and colleagues showed that unilateral damage (involving one hemisphere only) could results in similar symptoms. Their data also indicated the presence of other deficits such as an inability to orient the hand to certain angular targets, in the absence of problems with eye movements and spatial attention. This indicated that perhaps there may be some effector specificity in PPC. In other words perhaps there was separate subregions responsible for eye movements, hand movements and spatial attention within PPC.
Others then went on to propose that the primary deficit in these patients is a slowing of visual feedback used to guide hand movements.
Ultimately it was unclear whether PPC was doing purely visual processing, visuo-motor processing or visual feedback processing. Andersen made the point that it would be hard to make headway on this from lesion studies in humans as lesions tend to be large and take out extensive cortical regions, and what was needed was to look at the relatively small subsections of PPC.
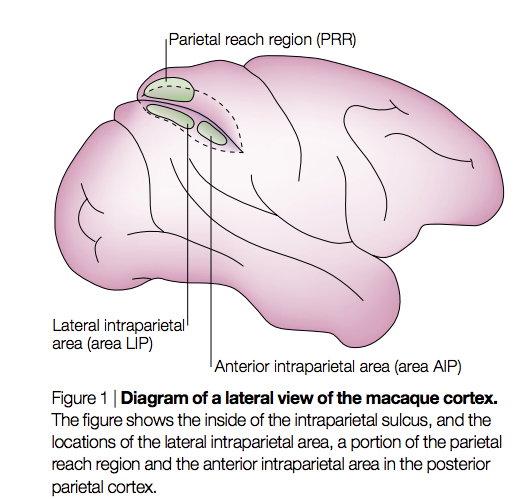
He went on to detail some animal work that made some headway into these issues. In summary they and others went on to show that non-human primate PPC has distinct subregions that encode these difference cognitive and motor functions. To summarise, a number of studies using reversible and irreversibly inactivation of small cortical regions demonstrated that with PPC, LIP is responsible for eye movements (saccades), posterior reaching region (PRR) for visually guided reaching, AIP for grasping movements of the hand (see figure).
However, there was the important question of whether non-human primate PPC and human PPC were organized in a similar manner. Functional MRI studies from a number of labs indicated some functional specialisation with PPC, but others showed that there was some multifunctional overlap between the regions involved in eye movements, reaching, grasping and attention. However, fMRI, despite providing some of the best spatial resolution possible for a non-invasive procedure, can only show us regions around 3mm apart. Transcranial magnetic stimulation (TMS) can sometimes spatially resolve things which fMRI cannot and Andersen discussed some data to this effect.
So the next logical step to address this question was to investigate intracranial recordings from human PPC. Andersen then went on to show videos of a patient with PPC electrodes implanted and how the signals from PPC can be used to control movements of a robotic arm with an impressive number of degrees of freedom in how it can move. He showed a patient who was asked to imagine making a movement and from this data they were able to decode the intended movement from his brain data in real time to move the robotic arm. The apparent benefits of using PPC signals, rather than primary motor cortex signals is that PPC seems to encode the goal of the movement, as well as the trajectory and importantly encodes information for bimanual movements. M1 on the other hand only relates to the contralateral limb.
Interestingly (for me) when the patient’s therapist manually moved his limb and he watched this movement, the movements of the robotic arm where impressively accurate. This is interesting as parietal cortex in the non-human primate contains mirror neurons, cells which respond to both the execution and the observation of an action. Andersen alluded to the idea that here, observation of one’s own limb was perhaps eliciting activity in parietal mirror neurons, and that it is this activity being picked up in PPC leading to the really impressively accurate robotic arm movements. When asked, Andersen said that at least initially, the observational PPC signals produce more accurate movements that the intentional PPC signals, which is fascinating to me. He said after a bit of training, the intentional signals get better as the patient learns how to interact with the arm. The patient was also able to manipulate the arm to make grasping actions. He showed his patient using the arm to grasp an apple, which was extremely impressive – grasping is notoriously hard to do without crushing the object in question.
At the end of it all, it still wasn't clear to me whether human PPC has functional specificity as the non-human primate PPC does, or whether there was some degree of multifunctionality. It seems that question might still be up for grabs.
Some questions of interest to us in the lab:
-Where do auditory-motor actions factor into this depiction of PPC? Is there a region in PPC that cares about auditory motor interactions of the hand (for example clapping) or articulators (such as speech)? Traditionally, dorsal (north) aspects of PPC are known to be associated with hand and arm movements, and more ventral (south) aspects with movements of the articulatory system. We also know that certain parts of macaque ventral parietal cortex respond to both execution of an action and hearing someone else making that action (auditory-motor mirror neurons). So where is the overlap or disctinction between effector specificity and feedback modality specificity in this?
- If observing an action is enough to produce a movement of the robotic arm, how do we go about preventing automatic imitation of the movements of others (echopraxia)?
- If cells in PPC are encoding vision of movement in the first and third person perspective, is there a role for PPC in linking the two?
I’ve missed all sorts of exciting things from the talk but you can’t remember it all. Anyway, it was very exciting.